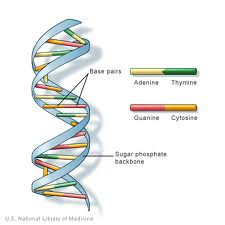
DNA (deoxyribonucleïnezuur) is de genetische informatie die in de kernen van de meeste organismen wordt aangetroffen. Het wordt gerangschikt in structuren genoemd chromosomen. De structuur van DNA werd eerst geà dentificeerd zoals het hebben van een ‘dubbel-helix’ structuur door Watson en Crick in 1953. DNA bestaat uit 4 basen: de purines, adenine (A) en guanine (G) en pyrimidines ,thymine (T) en cytosine (C). Deze vormen complementaire basenparen van A-T en G-C., DNA bevat ook een fosfaatgroep die met een deoxyribosesuiker wordt verbonden. De fosfaatgroep is via een fosfodiësterbinding aan de suiker gebonden. Mensen hebben 99,5% overeenkomsten met andere mensen in hun DNA.
structuur van DNA
DNA (deoxyribonucleïnezuur) is een keten van monomeren (zich herhalende eenheden) die “nucleotiden”worden genoemd. Een nucleotide bestaat uit: een 2` deoxyribose suiker (een vijf-koolstof pentose gelijkend op dat van ribose suiker gevonden in RNA., De chemische formule is C5H10O4), een fosfaatgroep (die een fosfodiësterbinding vormt: 2 deoxyribose-suikers verbinden) en een stikstofhoudende base (één uit A (Adenine), C ( Cytosine), g ( Guanine) of T (Thymine), die een zijketen vormt die vertakt wordt uit de 1′ koolstof van de 2` deoxyribose-suiker).
het gebied van de desoxyribosesuiker / fosfaatgroep wordt gezien als de “backbone”van DNA-strengen vanwege zijn structurele doel en de sequentie van basen draagt de gentische informatie. Om een double-stranded structuur van DNA te produceren, komen de interactie tussen aanvullende basissen voor., De complementaire basenparen in DNA interageren met elkaar via waterstofbindingen: A-T interacties bestaan uit 2 intermoleculaire waterstofbindingen, terwijl G-C interacties bestaan uit 3 intermoleculaire waterstofbindingen. Tussen deze basen zijn hydrofobe interacties bekend als Van der Waal krachten. Deze interactie vormt bruggen tussen twee kettingen van DNA, waarbij een dubbelstrengelde ‘ladder’ – gevormde structuur wordt gecreëerd. Elke bundel doet dienst als malplaatje voor andere in de replicatie van DNA., DNA wordt gekopieerd in mRNA (boodschappersRNA) dat de informatie van de originele malplaatjestreng van DNA draagt om bij eiwitsynthese te worden betrokken. Het proces van DNA dat in mRNA wordt gekopieerd wordt genoemd transcriptie. Het getranscribeerde mRNA wordt dan vertaald naar een polypeptide in een proces genoemd vertaling door tRNA.
in de dubbele helix van DNA liggen de strengen van de ruggengraat dichter bij elkaar aan de ene kant van de helix dan aan de andere. Dit leidt tot de vorming van grote en kleine groeven., De belangrijkste groef is veel breder dan de kleine groef en dit betekent dat specifieke DNA-eiwit interacties kunnen plaatsvinden op de belangrijkste groef toe te schrijven aan de ruggengraat niet in de weg. De specifieke nucleotiden die in de belangrijkste groef worden geconfronteerd zijn de N7 en C6 groepen van purines en de C4 en C5 groepen van pyrimidines, die waterstofionen van de aminozuren in het eiwit accepteren om waterstofbindingen te vormen.
door de dubbele spiraalvormige structuur van DNA worden de stikstofhoudende basen aan de binnenkant van de structuur gevonden, waardoor een hydrofoob interieur ontstaat., De negatieve lading van de fosfaatgroepen geeft de suiker-fosfaat backbone van DNA een negatieve lading, die nucleofielen afstoot, met inbegrip van water. Dit maakt DNA minder kwetsbaar voor nucleofiele aanval, dus DNA wordt beschouwd als een zeer stabiele molecule. DNA is veel stabieler dan RNA aangezien RNA slechts single-stranded is-de stikstofhoudende basissen worden verlaten blootgesteld aan aanval door nucleofielen aan één kant. in 1953, ondanks vele andere theorieën, ontdekten James Watson en Francis Crick de ware structuur van een dubbelstrengs DNA-molecuul als een ‘dubbele Helix’., Dit werd opgelost als gevolg van’ stick-and-ball ‘ -modellen die ze creëerden, samen met het werk van collega-wetenschappers Rosalind Franklin en Maurice Wilkins over röntgenkristallografie. De röntgendiffractiefoto ‘ s verkregen van DNA-vezels, toonden een unieke X-vorm, die een spiraalvormige structuur illustreert, hoewel ze een zich herhalende structuur van 3,4 Å uit elkaar per draai van de helix aangeven, wordt elke basis 36 graden gedraaid ten opzichte van de volgende. De diameter van de helix is 23,7 Å., Ze vonden dat de suiker-fosfaat backbone aan de buitenkant was en de basen zijn geplaatst aan de binnenkant van de helix.
bovenstaande informatie beschrijft de B-vorm van DNA. DNA wordt ook gevonden in a-en Z-vormen. Wanneer het DNA wordt uitgedroogd, kan de A-vorm worden waargenomen. Het is ook rechtshandig, maar er zijn 11 basen per beurt en de helix is breder. De diameter is 25,5 Å. Een ander verschil is dat de helling van de basenparen toeneemt met 18o, tot 19o van loodrecht op de spiraalas.
De Z-vorm verschilt veel meer omdat het een linkshandig dubbele helix is., Deze vorm wordt zelden gezien zonder de hulp van hoge zoutconcentraties. De bindingen zijn zigzagged als de bindingen afwisselend anti en syn zijn (terwijl A-en B-vormen alleen anti zijn). De Z-vorm is smaller, met een diameter van slechts 18,4 Å, maar er is een 3,8 Å stijging per basenpaar. Men denkt dat de overgangen tussen de B en Z vormen van DNA bij de regulatie van genregulatie kunnen worden betrokken.
B-DNA wordt het vaakst gezien in alle levensvormen, echter, A-spiraalvormige en Z-spiraalvormige structuren bestaan samen in cellen; d.w.z., het is heel gebruikelijk om een molecuul van B-DNA en Z-DNA waar te nemen , in een overwegend B-DNA bevestiging.
het DNA van de Indische muntjak, een Aziatisch hert, heeft de langste lengte (ongeveer 3 miljard nucleotiden) van alle bekende DNA-moleculen van andere organismen.
DNA is negatief geladen als gevolg van de negativley geladen fosfaationen in de suikerfosfaat backbone. Vandaar kan het voor gelelektroforese worden gebruikt om verschillende lengtes van DNA te identificeren., De negatieve lading van de backbone, samen met de OH-groepen op de deoxyribose suiker, betekent dat de backbone Hydrofillisch is aangezien water waterstofbanden met het kan vormen. Het centrum van de molecule van DNA is hydrophobic toe te schrijven aan het gebrek aan last in de basissen van DNA. De hydrofillische buiten-en hydrofobe binnenzijde van de DNA-molecule betekent dat het oplosbaar is in water.
replicatie
De dubbelstrengeling van DNA is belangrijk voor de “semi-conservatieve replicatie” methode van DNA-replicatie., In dit proces, wikkelt de helicase van enzym DNA de dubbele helix af door de waterstofbanden tussen de complementaire basissen op elke bundel te breken die de 2 afzonderlijke bundels openbaren. Op deze bundels zijn de onthulde basissen, die complementaire basissen op vrije nucleotiden aantrekken. De vrije nucleotiden worden met elkaar verbonden door de polymerase van enzym DNA. Deoxyribonucleotidetrifosfaat (dNTPs) wordt toegevoegd aan de 3′ hydroxylgroep op de groeiende bundel door de 5′ trifosfaatgroep op de inkomende dNTP in een esterificatiereactie., Het toetreden van nucleotiden vormt een nieuwe bundel van DNA die aan de andere dubbele bundel van DNA identiek is, aangezien het één van de originele bundels als malplaatje voor replicatie gebruikt. Elke dochter dubbele bundel van DNA wordt samengesteld uit een ouder bundel en een nieuw gesynthetiseerde bundel.
hoewel beide strengen in het DNA-molecuul van de ouders worden gekopieerd om identieke producten te vormen, worden de twee strengen op een enigszins andere manier van elkaar gekopieerd. Dit is toe te schrijven aan het feit dat DNA altijd in een 5′ Aan 3′ richting wordt gesynthetiseerd., De bundel 3′ aan 5′, die als belangrijke bundel wordt bekend, wordt gekopieerd onophoudelijk door de polymerase van DNA. De andere bundel wordt genoemd de achterblijvende bundel, aangezien het langzamer wordt gerepliceerd. Om de achterblijvende bundel te herhalen, worden de primers van RNA geplaatst op verscheidene punten langs de achterblijvende bundel door een enzym genoemd primase. De hiaten op de achterblijvende bundel tussen de inleidingen van RNA worden gerepliceerd door de polymerase van DNA, en de korte fragmenten van gerepliceerde DNA zijn genoemd geworden fragmenten van Okazaki. Nochtans, om de replicatie van de achterblijvende bundel te voltooien, moeten de primers van RNA door de opeenvolgingen van DNA worden vervangen., Een andere polymerase van DNA verwijdert de primers van RNA en synthetiseert de fragmenten van DNA om hen te vervangen. De Okazaki fragmenten en de RNA primer vervangingen zijn nog steeds niet samengevoegd, dus DNA ligase komt in een ligaten alle fragmenten van DNA samen.
De theorie van de semi-conservatieve replicatie bleek correct te zijn door het experiment Messelson-Stahl. In dit experiment werd E. coli gedurende een aantal generaties gekweekt in een medium dat 15-N bevat. De bacteriën werden toen overgebracht naar een medium dat 14-N. bevat nadat één DNA van de replicatiecyclus uit de bacteriën werd gehaald en werd gecentrifugeerd., De centrifugering scheidde het DNA door dichtheid, producerend één band wita h dichtheid tussen die van 15-N DNA en 14-N DNA. Dit toonde aan dat één bundel van de ouder (15-N) kwam en één bundel nieuw uit vrije nucleotiden (14-N) werd samengesteld.