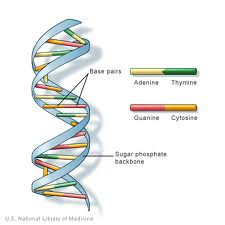
DNS (dezoxiribonukleinsav) a legtöbb szervezet magjaiban található genetikai információ. A kromoszómáknak nevezett struktúrákba van rendezve. A DNS szerkezetét először Watson és Crick 1953-ban azonosították “kettős spirál” szerkezettel. A DNS 4 bázisból áll: a purinokból, az adeninből (A) és a guaninból (G) és a pirimidinekből ,a timinból (T) és a citozinból (C). Ezek egymást kiegészítő a-T és G-C bázispárokat alkotnak., A DNS tartalmaz egy foszfátcsoportot is, amely dezoxiribóz cukorhoz kapcsolódik. A foszfátcsoport foszfodiészterkötéssel kapcsolódik a cukorhoz. Az emberek 99,5% – os hasonlóságot mutatnak más emberekkel a DNS-ben.
A DNS szerkezete
a DNS (dezoxiribonukleinsav) monomerek (ismétlődő egységek) lánca, úgynevezett “nukleotidok”. A nukleotid a következőkből áll: egy 2 ‘ dezoxiribózcukor (egy öt szén-pentóz, amely hasonló az RNS-ben található ribózcukoréhoz., Kémiai képlete C5H10O4), egy foszfátcsoport( amely foszfodiészterkötést képez: 2 dezoxiribózcukrot összeköt) és egy nitrogénbázis (az egyik a (adenin), C ( citozin), G ( guanin) vagy T (timin), amely a 2 “dezoxiribózcukor 1” szénéből elágazó oldalláncot képez).
a dezoxiribózcukor/foszfátcsoport régió szerkezeti célja miatt a DNS-szálak “gerincének” tekinthető, és a bázisok szekvenciája hordozza a gentikus információt. Annak érdekében, hogy kettős szálú DNS-struktúrát hozzon létre, kölcsönhatások lépnek fel a kiegészítő bázisok között., A DNS-ben lévő kiegészítő bázispárok hidrogénkötésekkel kölcsönhatásba lépnek egymással: az a-T kölcsönhatások 2 intermolekuláris hidrogénkötésből állnak, míg a G-C kölcsönhatások 3 intermolekuláris hidrogénkötésből állnak. Ezen bázisok között hidrofób kölcsönhatások vannak, amelyeket van der Waal erőknek neveznek. Ezek a kölcsönhatások hidakat képeznek két DNS-lánc között, így kettős szálú “létra” alakú szerkezetet hoznak létre. Mindegyik szál sablonként működik a másik DNS-replikációban., A DNS-t mRNS-be (messenger RNS-be) másolják, amely az eredeti DNS-sablon szálból származó információkat hordozza, hogy részt vegyen a fehérjeszintézisben. Az mRNS-be másolt DNS folyamatát transzkripciónak nevezik. Az átírt mRNS-t ezután egy polipeptidre fordítják a tRNA által készített fordításnak nevezett folyamatban.
a DNS kettős hélixben a gerinc szálai közelebb vannak egymáshoz a hélix egyik oldalán, mint a másik oldalon. Ez nagyobb és kisebb hornyok kialakulásához vezet., A fő horony sokkal szélesebb, mint a kisebb horony, ami azt jelenti, hogy a fő horonyon specifikus DNS-fehérje kölcsönhatások léphetnek fel, mivel a gerinc nem áll az úton. A fő horonyba kerülő specifikus nukleotidok a purinok N7 és C6 csoportjai, valamint a pirimidinek C4 és C5 csoportjai, amelyek a fehérje aminosavaiból hidrogénkötéseket képeznek.
a DNS kettős spirális szerkezete miatt a nitrogénbázisok a szerkezet belsejében találhatók, hidrofób belső teret képezve., A foszfátcsoportok negatív töltése negatív töltést ad a DNS cukor-foszfát gerincének, amely taszítja a nukleofileket, beleértve a vizet is. Ez a DNS-t kevésbé sebezhetővé teszi a nukleofil támadásokkal szemben, így a DNS-t nagyon stabil molekulának tekintik. A DNS sokkal stabilabb, mint az RNS, mivel az RNS csak egyszálú – a nitrogénbázisokat az egyik oldalon nukleofilok támadják meg.
1953-ban, sok más elmélet ellenére, James Watson és Francis Crick felfedezték a kettős szálú DNS-molekula valódi szerkezetét, hogy “kettős spirál” legyen., Ezt az általuk létrehozott “stick-and-ball” modellek eredményeként oldották meg, valamint Rosalind Franklin és Maurice Wilkins tudóstársainak röntgenkristályográfián végzett munkáját használták fel. A DNS-szálakból nyert röntgendiffrakciós fényképek egyedi X-alakot mutattak, amely egy spirális stuktúrát ábrázol, bár a hélix fordulónként 3,4 Å ismétlődő szerkezetet jeleztek, mindegyik alapot 36 fokkal elforgatják a következőtől. A hélix átmérője 23,7 Å., Megállapították, hogy a cukor-foszfát gerince kívülről van, a bázisok a hélix belsejében helyezkednek el.
a fenti információk leírták a DNS B-formáját. A DNS A – és Z-alakban is megtalálható. Amikor a DNS kiszárad, az A-forma megfigyelhető. Jobbkezes is, de fordulónként 11 bázis van, a hélix pedig szélesebb. Átmérője 25,5 Å. Egy másik különbség az, hogy az alappárok dőlése 18O-val, 19O-ra emelkedik a hélix tengelyre merőlegesen.
A Z-forma sokkal inkább különbözik, mivel Balkezes kettős spirál., Ezt a formát ritkán látják magas sókoncentráció nélkül. A kötések cikcakkban vannak, mivel a kötések váltakozó anti és syn (míg az A – és B-formák csak anti). A Z-forma keskenyebb, átmérője mindössze 18,4 Å, de bázispáronként 3,8 Å emelkedés van. Úgy gondolják, hogy a DNS B és Z formái közötti átmenetek szerepet játszhatnak a génszabályozás szabályozásában.
A B-DNS-t leggyakrabban minden életformában észlelik, azonban a-spirális és Z-spirális struktúrák a sejtekben együtt exszistálódnak; azaz, nagyon gyakori a B-DNS és a Z-DNS molekula megfigyelése egy túlnyomórészt B-DNS megerősítésben.
Az Ázsiai szarvasnak számító indiai muntjac DNS-e a leghosszabb hosszúságú (körülbelül 3 milliárd nukleotid) más szervezetek összes ismert DNS-molekulája között.
a DNS negatív töltésű a cukor-foszfát gerincben lévő negatív töltésű foszfátionok miatt. Ezért alkalmazható gél-elektroforézisre a DNS különböző hosszainak azonosítására., A gerinc negatív töltése a dezoxiribózcukor OH-csoportjaival együtt azt jelenti, hogy a gerinc hidrofil, mivel a víz hidrogénkötéseket képezhet vele. A DNS-molekula középpontja hidrofób a DNS-bázisok töltésének hiánya miatt. A DNS molekula hidrofil külső és hidrofób belső része azt jelenti, hogy vízben oldódik.
replikáció
a DNS kettős szálú jellege fontos a DNS-replikáció “félig konzervatív replikációs” módszeréhez., Ebben a folyamatban a DNS helikáz enzim feloldja a kettős hélixet azáltal, hogy megtöri a hidrogénkötéseket az egyes szálak kiegészítő bázisai között, feltárva a 2 különálló szálat. Ezeken a szálakon a feltárt bázisok, amelyek kiegészítő bázisokat vonzanak a szabad nukleotidokra. A szabad nukleotidokat egy enzim DNS-polimeráz köti össze. Dezoxiribonukleotid-trifoszfátot (dNTPs) adunk a növekvő szálon lévő 3′ hidroxilcsoporthoz az 5′ trifoszfátcsoporton keresztül a bejövő dNTP-n észterezési reakcióban., A nukleotidok összekapcsolása egy új DNS-szálat képez, amely megegyezik a DNS másik kettős szálával, mivel az eredeti szálak egyikét használja a replikáció sablonjaként. Minden lány kettős DNS-szál egy szülő szálból és egy újonnan szitézizált szálból áll.
annak ellenére, hogy a szülői DNS-molekula mindkét szálát átmásolják, hogy azonos termékeket képezzenek, a két szálat kissé eltérő módon másolják egymástól. Ez annak köszönhető, hogy a DNS-t mindig 5-3 irányban szintetizálják., A 3′ – 5 ‘ szál, a vezető szál néven ismert, DNS-polimeráz folyamatosan másolja. A másik szálat lemaradó szálnak nevezik, mivel lassabban replikálódik. A lemaradó szál replikálásához az RNS-alapozókat a lemaradó szál mentén több pontra helyezzük egy primáz nevű enzim segítségével. Az RNS-primerek közötti lemaradó szál réseit DNS-polimeráz replikálja, a replikált DNS rövid fragmenseit Okazaki fragmenseknek nevezik. A lemaradó szál replikációjának befejezéséhez azonban az RNS-alapozókat DNS-szekvenciákkal kell helyettesíteni., Egy másik DNS-polimeráz eltávolítja az RNS-primereket és DNS-fragmenseket szintetizál, hogy helyettesítse őket. Az Okazaki-fragmensek és az RNS-primer replacementek még mindig nem kapcsolódnak egymáshoz, így a DNS-ligáz egy ligátba kerül a DNS összes fragmentumával együtt.
a félig konzervatív replikáció elméletét a Messelson-Stahl kísérlet igazolta. Ebben a kísérletben az E. coli-t 15-N-t tartalmazó közegben termesztették több generáció számára. A baktériumokat ezután egy 14-N-t tartalmazó közegbe helyezték át, miután egy replikációs ciklus során DNS-t kivontak a baktériumokból és centrifugálták., A centrifugálás sűrűség szerint elválasztotta a DNS-t, egy sáv wita h sűrűséget eredményezett a 15-N DNS és a 14-N DNS között. Ez azt mutatta, hogy az egyik szál a szülőtől származik (15-N), az egyik szálat pedig újonnan szintetizálták szabad nukleotidokból (14-N).