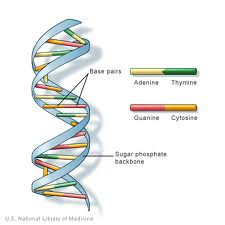
l’ADN (acide désoxyribonucléique) est l’information génétique trouvé dans les noyaux de la plupart des organismes. Il est organisé en structures appelées chromosomes. La structure de l’ADN a été identifiée pour la première fois comme ayant une structure en « double hélice » par Watson et Crick en 1953. L’ADN est composé de 4 bases: les purines, l’adénine (A) et la guanine (G) et les pyrimidines ,la thymine (T) et la cytosine (C). Ceux-ci forment des paires de bases complémentaires de A-T et G-C., L’ADN contient également un groupe phosphate relié à un sucre désoxyribose. Le groupe phosphate est attaché au sucre par une liaison phosphodiester. Les humains ont 99,5% de similitudes avec d’autres humains dans leur ADN.
Structure de l’ADN
l’ADN (acide désoxyribonucléique) est une chaîne de monomères (unités répétitives) appelé « nucléotides ». Un nucléotide est constitué de: un sucre désoxyribose 2` (Un pentose à cinq carbones similaire à celui du sucre ribose présent dans l’ARN., Sa formule chimique est C5H10O4), un groupe phosphate (qui forme une liaison phosphodiester: reliant 2 sucres désoxyribose ensemble) et une base azotée (une à partir de A (Adénine), C ( Cytosine), G ( Guanine) ou T (Thymine), qui forme une chaîne latérale ramifiée à partir du carbone 1′ du sucre désoxyribose 2`).
La région du groupe sucre/phosphate désoxyribose est considérée comme la « colonne vertébrale » des brins d’ADN en raison de son objectif structurel et la séquence de bases porte l’information gentique. Afin de produire une structure d’ADN double brin, des interactions se produisent entre les bases complémentaires., Les paires de bases complémentaires de l’ADN interagissent entre elles par l’intermédiaire de liaisons hydrogène: Les interactions A-T sont constituées de 2 liaisons hydrogène intermoléculaires, tandis que les interactions G-C sont constituées de 3 liaisons hydrogène intermoléculaires. Entre ces bases se trouvent des interactions hydrophobes connues sous le nom de forces de van der Waal. Ces interactions forment des ponts entre deux chaînes d’ADN, créant ainsi une structure en forme d’échelle à double brin. Chaque brin agit comme un modèle pour l’autre dans la réplication de l’ADN., L’ADN est copié dans l’ARNm (ARN messager) qui porte l’information du brin original de modèle d’ADN pour être impliqué dans la synthèse de protéine. Le processus de copie de l’ADN dans l’ARNm est appelé transcription. L’ARNm transcrit est ensuite traduit en un polypeptide dans un processus appelé traduction par ARNt.
Dans la double hélice d’ADN, les brins de l’épine dorsale sont plus rapprochés d’un côté de l’hélice que de l’autre. Cela conduit à la formation de rainures majeures et mineures., Le sillon majeur est beaucoup plus large que le sillon mineur et cela signifie que des interactions ADN-protéine spécifiques peuvent avoir lieu sur le sillon majeur en raison de l’épine dorsale ne pas être de la manière. Les nucléotides spécifiques qui font face à la rainure principale sont les groupes N7 et C6 des purines et les groupes C4 et C5 des pyrimidines, qui acceptent les ions hydrogène des acides aminés de la protéine pour former des liaisons hydrogène.
En raison de la double structure hélicoïdale de l’ADN, les bases azotées se trouvent à l’intérieur de la structure, formant un intérieur hydrophobe., La charge négative des groupes phosphate donne à l’épine dorsale sucre-phosphate de l’ADN une charge négative, qui repousse les nucléophiles, y compris l’eau. Cela rend l’ADN moins vulnérable aux attaques nucléophiles, donc l’ADN est considéré comme une molécule très stable. L’ADN est beaucoup plus stable que l’ARN puisque l’ARN n’est qu’un brin-les bases azotées sont exposées à l’attaque par les nucléophiles d’un côté.
En 1953, malgré de nombreuses autres théories, James Watson et Francis Crick ont découvert que la véritable structure d’une molécule d’ADN double brin était une « Double hélice »., Cela a été résolu grâce aux modèles « stick-and-ball » qu’ils ont créés, ainsi qu’à l’utilisation des travaux de leurs collègues scientifiques Rosalind Franklin et Maurice Wilkins sur la cristallographie aux rayons X. Les photographies de diffraction des rayons X obtenues à partir de fibres d’ADN ont montré une forme en X unique, qui illustre une structure hélicoïdale, bien qu’elles indiquent une structure répétitive de 3,4 Å par tour de l’hélice, chaque base est tournée de 36 degrés par rapport à la suivante. Le diamètre de l’hélice est de 23,7 Å., Ils ont constaté que le squelette sucre-phosphate était à l’extérieur, et les bases sont positionnés à l’intérieur de l’hélice.
Les informations ci-dessus décrivent la forme B de l’ADN. L’ADN se trouve également sous les formes A et Z. Lorsque l’ADN se déshydrate, la forme peut être observée. Il est également droitier, mais il y a 11 bases par tour et l’hélice est plus large. Le diamètre est de 25,5 Å. Une autre différence est que l’inclinaison des paires de bases augmente de 18o, à 19o de perpendiculaire à l’axe de l’hélice.
La forme Z diffère beaucoup plus car il s’agit d’une double hélice gauchère., Cette forme est rarement vue sans l’aide de fortes concentrations de sel. Les liaisons sont zigzaguées car les liaisons alternent anti et syn (alors que les formes A et B ne sont anti que). La forme Z est plus étroite, ayant un diamètre de seulement 18,4 Å, mais il y a une élévation de 3,8 Å par paire de base. On pense que les transitions entre les formes B et Z de l’ADN peuvent être impliquées dans la régulation de la régulation des gènes.
L’ADN B est le plus souvent observé dans toutes les formes de vie, cependant, les structures A-hélicoïdales et Z-hélicoïdales coexistent dans les cellules; c’est-à-dire, il est très courant d’observer une molécule d’ADN-B et d’ADN-Z , dans une confirmation d’ADN-B prédominante.
L’ADN du muntjac indien qui est un cerf asiatique a la plus longue longueur (environ 3 milliards de nucléotides) parmi toutes les molécules d’ADN connues d’autres organismes.
L’ADN est chargé négativement en raison des ions phosphates chargés négativement dans l’épine dorsale sucre-phosphate. Par conséquent, il peut être utilisé pour l’électrophorèse sur gel pour identifier différentes longueurs d’ADN., La charge négative de l’épine dorsale, ainsi que les groupes OH sur le sucre désoxyribose, signifie que l’épine dorsale est hydrophile car l’eau peut former des liaisons hydrogène avec elle. Le centre de la molécule d’ADN est hydrophobe en raison de l’absence de charge dans les bases de l’ADN. L’intérieur hydrophile externe et hydrophobe de la molécule d’ADN signifie qu’elle est soluble dans l’eau.
Réplication
La nature double brin de l’ADN est importante pour la méthode de « réplication semi-conservatrice » de la réplication de l’ADN., Dans ce processus, l’enzyme ADN hélicase déroule la double hélice en brisant les liaisons hydrogène entre les bases complémentaires de chaque brin révélant les 2 brins séparés. Sur ces brins sont les bases révélées, qui attirent les bases complémentaires sur les nucléotides libres. Les nucléotides libres sont réunis par une enzyme ADN polymérase. Le triphosphate de désoxyribonucléotide (dNTPs) est ajouté sur le groupe hydroxyle 3′ sur le brin en croissance à travers le groupe triphosphate 5′ sur le dNTP entrant dans une réaction d’estérification., L’assemblage de nucléotides forme un nouveau brin d’ADN qui est identique à l’autre double brin d’ADN, car il utilise l’un des brins d’origine comme modèle de réplication. Chaque double brin d’ADN fille est composé d’un brin parent et d’un brin nouvellement sythesisé.
Même si les deux brins de la molécule d’ADN parentale sont copiés pour former des produits identiques, les deux brins sont copiés d’une manière légèrement différente l’un de l’autre. Cela est dû au fait que l’ADN est toujours synthétisé dans une direction 5′ à 3′., Le brin 3′ à 5′, connu sous le nom de brin principal, est copié en continu par l’ADN polymérase. L’autre brin est appelé le brin retardant, car il est répliqué plus lentement. Pour répliquer le brin en retard, des amorces d’ARN sont placées sur plusieurs points le long du brin en retard par une enzyme appelée primase. Les lacunes sur le brin en retard entre les amorces d’ARN sont répliquées par l’ADN polymérase, et les fragments courts d’ADN répliqué sont connus sous le nom de fragments d’Okazaki. Cependant, afin d’achever la réplication du brin en retard, les amorces d’ARN doivent être remplacées par des séquences d’ADN., Une autre ADN polymérase enlève les amorces d’ARN et synthétise des fragments d’ADN pour les remplacer. Les fragments d’Okazaki et les remplacements d’amorce d’ARN ne sont toujours pas joints, ainsi la ligase d’ADN vient dans un ligate tous les fragments d’ADN ensemble.
La théorie de la réplication semi-conservatrice s’est avérée correcte par l’expérience de Messelson-Stahl. Dans cette expérience, E. coli ont été cultivés dans un milieu contenant 15-N pendant un certain nombre de générations. Les bactéries ont ensuite été transférées dans un milieu contenant du 14-N. Après un cycle de réplication, l’ADN a été extrait des bactéries et centrifugé., La centrifugation a séparé l’ADN par densité, produisant une densité wita h de bande entre celle de l’ADN 15-N et de l’ADN 14-N. Cela a montré qu’un brin provenait du parent (15-N) et qu’un brin était nouvellement synthétisé à partir de nucléotides libres (14-N).