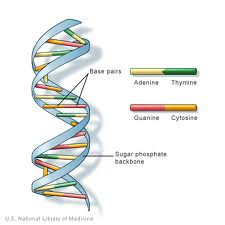
DNA (deoxyribonukleové kyseliny) je genetické informace nalezen v jádrech většiny organismů. Je uspořádán do struktur nazývaných chromozomy. Struktura DNA byla poprvé identifikována jako struktura „dvojité šroubovice“ Watsonem a Crickem v roce 1953. DNA se skládá ze 4 bází: purinů, adenin (a) a guaninu (G) a pyrimidinů ,thyminu (T) a cytosinu (C). Ty tvoří komplementární základní páry A-T A G-C., DNA také obsahuje fosfátovou skupinu spojenou s deoxyribózovým cukrem. Fosfátová skupina je připojena k cukru prostřednictvím fosfodiesterové vazby. Lidé mají 99,5% podobnosti s ostatními lidmi v jejich DNA.
struktura DNA
DNA (deoxyribonukleová kyselina) je řetězec monomerů (opakujících se jednotek) nazývaný „nukleotidy“. Nukleotid se skládá z: 2` deoxyribose cukru (pět uhlíku pentóza podobné ribóza cukr nalézt v RNA., Jeho chemický vzorec je C5H10O4), fosfátové skupiny (které tvoří phosphodiester bond: připojení 2 deoxyribose cukry dohromady) a dusíkaté báze (jeden z A (Adenin), C ( Cytosin), G ( Guanin) a T (Thymin), které tvoří postranní řetězec odbočení od 1′ uhlíku 2` deoxyribose cukru).
deoxyribose cukru/fosfátová skupina region je považován za „páteř“ vlákna DNA v důsledku své strukturální účel a pořadí bází nese genetická porucha, informace. Za účelem vytvoření dvouvláknové struktury DNA dochází k interakcím mezi komplementárními bázemi., Komplementární páry bází v DNA vzájemně interagují prostřednictvím vodíkových vazeb: interakce A-T se skládají ze 2 intermolekulárních vodíkových vazeb, zatímco interakce G-C se skládají ze 3 intermolekulárních vodíkových vazeb. Mezi těmito základnami jsou hydrofobní interakce známé jako Van der Waalovy síly. Tyto interakce tvoří mosty mezi dvěma řetězci DNA, čímž vzniká dvouvláknová struktura ve tvaru „žebříku“. Každý pramen působí jako šablona pro druhou v replikaci DNA., DNA je zkopírována do mRNA (messenger RNA), který přenáší informace z původního templátu DNA strand být zapojen do syntézy bílkovin. Proces kopírování DNA do mRNA se nazývá transkripce. Transkribovaná mRNA je pak přeložena do polypeptidu v procesu zvaném translation by tRNA.
v DNA dvojité šroubovice jsou prameny páteře blíže k sobě na jedné straně šroubovice, než jsou na druhé straně. To vede k tvorbě hlavních a menších drážek., Hlavní drážka je mnohem širší než menší drážka, což znamená, že specifické interakce DNA-bílkovin mohou probíhat na hlavní drážce kvůli tomu, že páteř není v cestě. Specifické nukleotidy, které obličej do hlavní drážky jsou N7 a C6 skupiny purinů a C4 a C5 skupiny pyrimidinů, které přijímají vodíkové ionty z aminokyselin v proteinu tvořit vodíkové vazby.
vzhledem k dvojité spirálové struktuře DNA se dusíkaté báze nacházejí na vnitřní straně struktury a vytvářejí hydrofobní interiér., Negativní náboj z fosfátových skupin dává cukr-fosfátová páteř DNA negativní náboj, který odpuzuje nucleophiles, včetně vody. Díky tomu je DNA méně zranitelná vůči nukleofilnímu útoku, takže DNA je považována za velmi stabilní molekulu. DNA je mnohem stabilnější než RNA, protože RNA je pouze jednovláknová-dusíkaté báze jsou na jedné straně vystaveny útoku nukleofilů.
v roce 1953, navzdory mnoha dalším teoriím, James Watson a Francis Crick objevili skutečnou strukturu dvouvláknové molekuly DNA jako „dvojitou šroubovice“., To byl vyřešen v důsledku ‚stick-a-ball‘ modely, které vytvořili, spolu s využitím práce kolegů vědců Rosalinda Franklinová a Maurice Wilkins na X-ray krystalografie. X-ray difrakce fotografie získané z DNA vláken, zobrazí unikátní X-tvar, který znázorňuje helikální strukturu, i když uvedly, opakující se struktury 3.4 Å od sebe na přelomu šroubovice, každá základna se otáčí 36 stupňů od vedle. Průměr šroubovice je 23,7 Å., Zjistili, že kostra cukru a fosfátu byla na vnější straně a základny jsou umístěny na vnitřní straně šroubovice.
výše uvedené informace popsaly B-formu DNA. DNA se také nachází ve formách A – A Z. Když se dna dehydratuje, lze pozorovat formu A. Je také pravák, ale na otočení je 11 základen a šroubovice je širší. Průměr je 25,5 Å. Dalším rozdílem je, že náklon základních párů se zvyšuje o 18o, na 19O od kolmice k ose šroubovice.
Z-forma se liší mnohem více, protože se jedná o levotočivou dvojitou šroubovice., Tato forma je zřídka vidět bez pomoci vysokých koncentrací soli. Vazby jsou klikaté, protože vazby se střídají proti a syn(zatímco formy A A B jsou pouze proti). Z-forma je užší, má průměr pouze 18,4 Å, ale na základní pár je 3,8 Å. Předpokládá se, že přechody mezi B A Z formy DNA mohou být zapojeny do regulace genové regulace.
B-DNA se nejčastěji vyskytuje ve všech formách života, avšak a-spirálové a z-spirálové struktury spolu exsistují v buňkách; tj., je velmi běžné pozorovat molekulu B-DNA A Z-DNA v převážně B-DNA potvrzení.
DNA Indického muntjac, což je Asijský jelen má nejdelší délku (přibližně 3 miliardy nukleotidů) mezi všechny známé molekuly DNA jiných organismů.
DNA je záporně nabitá v důsledku záporně nabitých fosfátových iontů v páteři cukru a fosfátu. Proto může být použita pro gelovou elektroforézu k identifikaci různých délek DNA., Záporný náboj páteře spolu s OH-skupinami na deoxyribózovém cukru znamená, že páteř je hydrofilní, protože voda s ním může tvořit vodíkové vazby. Střed molekuly DNA je hydrofobní kvůli nedostatku náboje v DNA bázích. Hydrofilní vnější a hydrofobní vnitřek molekuly DNA znamená, že je rozpustný ve vodě.
replikace
dvojvláknová povaha DNA je důležitá pro metodu „Polo-konzervativní replikace“ replikace DNA., V tomto procesu, enzym DNA helicase rozplete dvoušroubovice rozbitím vodíkových vazeb mezi komplementárními bázemi na každý pramen odhalující 2 oddělené prameny. Na těchto pramenech jsou odhalené báze, které přitahují komplementární báze na volných nukleotidech. Volné nukleotidy jsou spojeny enzymem DNA polymerázou. Deoxyribonucleotide trifosfátu (dntp) jsou přidány na 3′ hydroxylové skupiny na rostoucí pramen přes 5′ – trifosfát skupiny na příchozí dNTP v esterifikace reakce., Spojení nukleotidů tvoří nový řetězec DNA, který je totožný s druhým dvojitým pramenem DNA, protože používá jeden z původních pramenů jako šablonu pro replikaci. Každý dceřiný dvojitý řetězec DNA je tvořen mateřským pramenem a nově sythesizovaným pramenem.
přestože jsou oba prameny v molekule rodičovské DNA kopírovány tak, aby vytvářely identické produkty, jsou oba prameny kopírovány mírně odlišným způsobem od sebe. To je způsobeno skutečností, že DNA je vždy syntetizována ve směru 5 „až 3“., Vlákno 3 ‚až 5‘, známé jako přední pramen, je kopírováno kontinuálně DNA polymerázou. Druhý pramen se nazývá zaostávající pramen, protože se replikuje pomaleji. K replikaci zaostávající strand, RNA primery jsou umístěny na několika místech podél zaostávající strand pomocí enzymu zvaného primase. Mezery na zaostávající strand mezi RNA primery jsou replikovány pomocí DNA polymerázy a krátké fragmenty replikované DNA jsou známé jako Okazaki fragmenty. Aby však bylo možné dokončit replikaci zaostávajícího pramene, musí být primery RNA nahrazeny sekvencemi DNA., Další DNA polymeráza odstraňuje primery RNA a syntetizuje fragmenty DNA, aby je nahradila. Fragmenty Okazaki a náhrady primeru RNA stále nejsou spojeny, takže dna ligáza přichází v ligátech všechny fragmenty DNA dohromady.
teorie semi-konzervativní replikace byla prokázána jako správná experimentem Messelson-Stahl. V tomto experimentu byla E. coli pěstována v médiu obsahujícím 15-N po řadu generací. Bakterie byly poté přeneseny do média obsahujícího 14-N. po jednom replikačním cyklu byla DNA extrahována z bakterií a centrifugována., Centrifugace oddělila DNA hustotou a vytvořila jednu hustotu pásma wita h mezi hustotou 15-N DNA a 14-N DNA. To ukázalo, že jeden pramen pochází od rodiče (15-N) A jeden pramen byl nově syntetizován z volných nukleotidů (14-N).