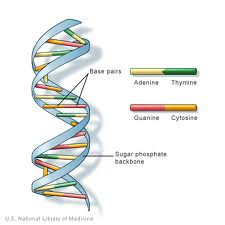
DNA (deoksiribonukleiinihappo) on geneettinen tieto löytyy ytimet useimmat organismit. Se on järjestetty kromosomeiksi kutsuttuihin rakenteisiin. DNA: n rakenne tunnistettiin ensimmäisen kerran Watsonin ja Crickin ”kaksoiskierteiseksi” rakenteeksi vuonna 1953. DNA koostuu 4 emäkset: puriinit, adeniini (A) ja guaniini (G) ja pyrimidines ,tymiini (T) ja sytosiini (C). Nämä muodostavat A-T: n ja G-C: n toisiaan täydentäviä emäspareja., DNA sisältää myös fosfaattiryhmän, joka on yhteydessä deoksiriboosisokeriin. Fosfaattiryhmä kiinnittyy sokeriin fosfodiesterisidoksen kautta. Ihmisillä on DNA: ssaan 99,5% yhtäläisyyksiä muihin ihmisiin.
DNA Rakenne
DNA (deoksiribonukleiinihappo) on ketjun monomeerien (toistuvat yksiköt) nimeltään ”nukleotidien”. Nukleotidi koostuu: 2` deoksiriboosisokerista (RNA: ssa esiintyvän riboosisokerin kaltainen viiden hiilen pentoosi., Sen kemiallinen kaava on C5H10O4), fosfaatti ryhmä (joka muodostaa phosphodiester sidos: yhdistää 2 deoxyribose sokereita yhdessä) ja typpipitoisia base (yksi A (Adeniini), C ( Sytosiini), G ( Guaniini) ja T (Tymiini), joka muodostaa sivuketjun aluevaltaus 1′ hiili 2` deoxyribose sokeria).
deoxyribose sokeria/fosfaatti-ryhmän alueella pidetään ’selkäranka’ DNA-säikeet, koska sen rakenteellinen tarkoitus ja järjestys emäkset kantaa gentic tietoja. Kaksijuosteisen DNA-rakenteen tuottamiseksi syntyy vuorovaikutuksia toisiaan täydentävien emästen välillä., Täydentävä emäsparin DNA vuorovaikutuksessa toistensa kanssa kautta vetysidokset: Klo vuorovaikutukset koostuvat 2 molekyylien välisten vetysidosten, kun taas G-C vuorovaikutukset koostuvat 3 molekyylien välisten vetysidosten. Näiden emästen välissä on hydrofobisia vuorovaikutuksia, jotka tunnetaan nimellä van der Waalin voimat. Nämä vuorovaikutussuhteet muodostavat siltoja kahden DNA-ketjut, luoden double-stranded ’tikkaat-muotoinen rakenne. Kukin juoste toimii mallina toiselle DNA-replikaatiossa., DNA kopioidaan mRNA (messenger RNA), joka kuljettaa tietoja alkuperäisen DNA: n mallin strand olla mukana proteiinisynteesiä. DNA: n kopioimista mRNA: ksi kutsutaan transkriptioksi. Litteroitu mRNA on sitten käännetty polypeptidi, prosessi nimeltä käännös tRNA.
DNA: n kaksoiskierteessä selkärangan säikeet ovat lähempänä toisiaan helixin toisella puolella kuin toisella. Tämä johtaa suurten ja pienten urien muodostumiseen., Merkittävä ura on paljon laajempi kuin pieni ura ja tämä tarkoittaa, että erityisiä DNA-proteiini vuorovaikutus voi tapahtua merkittäviä ura, koska selkäranka ei ole tiellä. Erityisiä nukleotidin, että kasvot osaksi suuret ura ovat N7 ja C6 ryhmien puriinit ja C4 ja C5 ryhmien pyrimidines, jotka hyväksyvät vetyioneja aminohappoja proteiini muodostaa vetysidoksia.
Koska double helix DNA: n rakenteen, ja typpipitoiset emäkset ovat löysi sisäpuolella rakenteen, muodostaen hydrofobisen sisustus., Negatiivinen varaus peräisin fosfaatti-ryhmien antaa sokeri-fosfaatti selkäranka DNA-negatiivinen varaus, joka hylkii nucleophiles, mukaan lukien veden. Tämä tekee DNA vähemmän alttiita nukleofiiliset hyökkäys, jolloin DNA: ta pidetään erittäin vakaa molekyyli. DNA on paljon vakaampi kuin RNA, koska RNA on vain yksijuosteinen – typpiemäkset jäävät alttiiksi nukleofiilien hyökkäykselle toisella puolella.
Vuonna 1953, vaikka monia muita teorioita, James Watson ja Francis Crick löysi todellisen rakenteen kaksinkertaisen pulaan DNA-molekyylin olevan ’Double Helix’., Tämä oli ratkaistu seurauksena keppi-ja-pallo mallit, he loivat yhdessä työn hyödyntämiseen tutkijoita Rosalind Franklin ja Maurice Wilkins X-ray crystallography. X-ray diffraction valokuvia saatu DNA: ta, kuituja, näkyy ainutlaatuinen X-muoto, joka osoittaa, kierteiset rakennetta, vaikka he ilmoittivat, toistuva rakenne 3.4 Å toisistaan vuorollaan helix, jokainen pohja on 36 astetta kääntää seuraavaan. Helixin halkaisija on 23,7 Å., He havaitsivat, että sokeri-fosfaattirunko oli ulkopuolella ja emäkset on sijoitettu helixin sisäpuolelle.
yllä olevat tiedot kuvasivat DNA: n B-muotoa. DNA: ta on myös A – ja Z-muodoissa. Kun DNA: sta tulee nestehukkaa, voidaan havaita a-muoto. Se on myös oikeakätinen, mutta on 11 emäkset vuorollaan ja helix on laajempi. Halkaisija on 25,5 Å. Toinen ero on, että kallistus base paria kasvaa 18o, että 19o päässä kohtisuoraan helix-akselilla.
Z-muoto eroaa paljon enemmän, koska se on vasenkätinen kaksoiskierre., Tämä muoto näkyy harvoin ilman korkeiden suolapitoisuuksien apua. Joukkovelkakirjat ovat zigzagged kuin joukkolainat ovat vuorotellen anti-ja syn (taas A – ja B-muodot ovat anti vain). Z-muoto on kapeampi, sen halkaisija on vain 18,4 Å, mutta perusparia kohden on 3,8 Å: n nousu. Ajatellaan, että siirtymät DNA: n B-ja Z-muotojen välillä voivat olla mukana geenien säätelyn säätelyssä.
B-DNA: ta on yleisimmin nähty kaikissa elämän muodoissa, kuitenkin, A-kierteiset ja Z-kierteiset rakenteet co-exsist soluissa; ts., on hyvin yleistä havaita B-DNA: n ja Z-DNA: n molekyyli pääasiassa B-DNA: n konfirmaatiossa.
DNA muntjakki, joka on Aasian hirvi on pisin pituus (noin 3 miljardia nukleotidia) joukossa kaikki tunnetut DNA-molekyylejä ja muita organismeja.
DNA on negatiivisesti varautunut sokeri-fosfaattirungon negativley-varautuneiden fosfaatti-ionien vuoksi. Siksi sitä voidaan käyttää geelielektroforeesissa DNA: n eri pituuksien tunnistamiseen., Selkärangan negatiivinen varaus yhdessä deoksiriboosisokerin OH-ryhmien kanssa tarkoittaa, että selkäranka on hydrofiilinen, koska vesi voi muodostaa sen kanssa vetysidoksia. DNA-molekyylin keskus on hydrofobinen, koska DNA-emäksissä ei ole latausta. DNA-molekyylin hydrofiilinen ulompi ja hydrofobinen sisempi tarkoittaa, että se liukenee veteen.
Replikaatio
double-stranded luonne DNA on tärkeää ”osittain konservatiivinen replikointi” menetelmä DNA: n replikaatio., Tässä prosessissa, entsyymi DNA helicase aueta double helix rikkomalla vetysidokset välillä täydentäviä perustaa kunkin lohkon paljastaen 2 erillinen osa. Nämä säikeet ovat paljasti, emäkset, jotka houkuttelevat toisiaan täydentäviä perustaa vapaita nukleotideja. Vapaita nukleotideja yhdistää DNA-polymeraasientsyymi. Deoxyribonucleotide trifosfaatiksi (dNTPs) lisätään päälle 3′ hydroksyyli-ryhmän kasvava lohkon läpi 5′ trifosfaatiksi ryhmän saapuvat dNTP vuonna esteröimällä reaktio., Liittymällä nukleotidien muodostaa uutta DNA: ta, joka on identtinen muiden double strand DNA, koska se käyttää yksi alkuperäinen osa mallina replikointi. Jokainen tytär kaksinkertainen DNA-juoste koostuu vanhemmasta juosteesta ja vastaperustetusta sythesisoidusta juosteesta.
vaikka molemmat säikeet vanhempien DNA-molekyyli on kopioitu muodostaa samanlaisia tuotteita, kaksi osaa on kopioitu hieman eri tavalla toisistaan. Tämä johtuu siitä, että DNA: ta syntetisoidaan aina 5-3-suunnassa., Johtavana juosteena tunnettu 3′ – 5 ’ – juoste kopioidaan jatkuvasti DNA-polymeraasilla. Toista juostetta kutsutaan lagging strandiksi, koska se toistuu hitaammin. Jäljitellä jäljessä strand, RNA-alukkeet ovat sijoitettu useita kohtia pitkin jäljessä strand nimisen entsyymin primase. Aukkoja jäljessä strand välillä RNA-alukkeet ovat monistaa DNA-polymeraasi, ja lyhyitä katkelmia monistaa DNA: ta tunnetaan Okazaki palasia. Kuitenkin, jotta täydellinen replikointi jäljessä strand, RNA-alukkeet on korvattava DNA-sekvenssit., Toinen DNA-polymeraasi poistaa RNA-alkulähteet ja syntetisoi niiden tilalle DNA-fragmentteja. Okazaki-sirpaleet ja RNA-pohjustuskorvikkeet eivät ole vielä liittyneet toisiinsa, joten DNA-ligaasi tulee ligaattina kaikki DNA: n sirpaleet yhteen.
Semikonservatiivisen replikaation teoria osoittautui oikeaksi Messelson-Stahlin kokeessa. Tässä kokeessa kolibakteeria kasvatettiin 15-N sisältävässä väliaineessa useiden sukupolvien ajan. Bakteerit olivat sitten siirretään väliaineeseen, jossa 14-N. Kun yksi replikointi sykli DNA uutettiin bakteerit ja sentrifugoidaan., Se sentrifugointi erotti DNA tiheys, tuottavat yksi bändi wita s tiheys välillä 15-N DNA-ja 14-N DNA: ta. Tämä osoitti, että yksi säie tuli vanhemman (15-N) ja yksi osa oli hiljattain syntetisoitu ilmainen nukleotidit (14-N).